การหมุนรอบแกนของมัน การหมุนรอบดวงอาทิตย์ของเรา โลกหมุนด้วยความเร็วเท่าใด
ตั้งแต่สมัยโบราณ ผู้คนสนใจว่าเหตุใดกลางคืนจึงหลีกทางให้กลางวัน ฤดูหนาวในฤดูใบไม้ผลิ และฤดูร้อนในฤดูใบไม้ร่วง ต่อมาเมื่อครั้งแรก...
ลีเซนโก แอนนา
เรียงความชีววิทยาให้คำจำกัดความของพันธุศาสตร์ ขั้นตอนการพัฒนาวิทยาศาสตร์นี้ และความสำคัญของวิทยาศาสตร์ต่อชีวิตมนุษย์
พันธุศาสตร์เป็นหนึ่งในสาขาวิชาหลักที่น่าสนใจที่สุดและในเวลาเดียวกันก็ซับซ้อนของวิทยาศาสตร์ธรรมชาติสมัยใหม่ สถานที่ทางพันธุศาสตร์ในสาขาวิทยาศาสตร์ชีวภาพและความสนใจเป็นพิเศษนั้นถูกกำหนดโดยข้อเท็จจริงที่ว่ามันศึกษาคุณสมบัติพื้นฐานของสิ่งมีชีวิต ได้แก่ พันธุกรรมและความแปรปรวน
จากการทดลองมากมายในสาขาอณูพันธุศาสตร์ การออกแบบที่ชาญฉลาดและการดำเนินการที่ประณีต ชีววิทยาสมัยใหม่จึงได้รับการเสริมสมรรถนะด้วยการค้นพบพื้นฐานสองประการ ซึ่งได้สะท้อนให้เห็นอย่างกว้างขวางในพันธุศาสตร์ของมนุษย์ และบางส่วนได้ดำเนินการในเซลล์ของมนุษย์ สิ่งนี้แสดงให้เห็นถึงความเชื่อมโยงที่แยกไม่ออกระหว่างความสำเร็จของพันธุกรรมมนุษย์กับความสำเร็จของชีววิทยาสมัยใหม่ ซึ่งมีความเชื่อมโยงกับพันธุกรรมมากขึ้นเรื่อยๆ
ประการแรกคือความสามารถในการทำงานร่วมกับยีนที่แยกได้ ได้มาจากการแยกยีนในรูปแบบบริสุทธิ์แล้วสังเคราะห์ ความสำคัญของการค้นพบนี้เป็นเรื่องยากที่จะประเมินค่าสูงไป สิ่งสำคัญคือต้องเน้นว่ามีการใช้วิธีการต่างๆ ในการสังเคราะห์ยีน เช่น มีทางเลือกอยู่แล้วเมื่อพูดถึงกลไกที่ซับซ้อนเช่นนี้ในฐานะบุคคล
ความสำเร็จประการที่สองเป็นการพิสูจน์ถึงการรวมข้อมูลแปลกปลอมไว้ในจีโนม ตลอดจนการทำงานของมันในเซลล์ของสัตว์และมนุษย์ชั้นสูง วัสดุสำหรับการค้นพบนี้สะสมมาจากวิธีการทดลองต่างๆ ก่อนอื่นนี่เป็นการศึกษาจำนวนมากในสาขาทฤษฎีไวรัสและพันธุศาสตร์ของการเกิดขึ้นของเนื้องอกมะเร็งรวมถึงการตรวจหาการสังเคราะห์ DNA บนเมทริกซ์ RNA นอกจากนี้การทดลองที่มีการถ่ายทอดคำพยากรณ์ซึ่งกระตุ้นโดยแนวคิดทางพันธุวิศวกรรมได้ยืนยันความเป็นไปได้ของการทำงานของยีนของสิ่งมีชีวิตอย่างง่ายในเซลล์ของสัตว์เลี้ยงลูกด้วยนมรวมถึงเซลล์ของมนุษย์ด้วย
หากไม่มีการพูดเกินจริง เราสามารถพูดได้ว่า นอกจากอณูพันธุศาสตร์แล้ว พันธุศาสตร์มนุษย์ถือเป็นสาขาหนึ่งของพันธุศาสตร์ที่ก้าวหน้าที่สุดโดยทั่วไป งานวิจัยของเธอขยายตั้งแต่ระดับชีวเคมีไปจนถึงระดับประชากร โดยผสมผสานระดับเซลล์และสิ่งมีชีวิต
แต่ให้เราพิจารณาประวัติความเป็นมาของการพัฒนาพันธุกรรมแยกกัน
ขั้นตอนหลักของการพัฒนาพันธุศาสตร์
ต้นกำเนิดของพันธุกรรมก็เหมือนกับวิทยาศาสตร์อื่นๆ ที่ควรถูกค้นหาในทางปฏิบัติ พันธุศาสตร์เกิดขึ้นจากการเพาะพันธุ์สัตว์เลี้ยงและการเพาะปลูกพืชตลอดจนการพัฒนายา เนื่องจากมนุษย์เริ่มใช้การผสมข้ามระหว่างสัตว์และพืช เขาต้องเผชิญกับความจริงที่ว่าคุณสมบัติและลักษณะของลูกหลานนั้นขึ้นอยู่กับคุณสมบัติของผู้ปกครองที่ได้รับเลือกให้ข้าม ด้วยการเลือกและข้ามลูกหลานที่ดีที่สุดจากรุ่นสู่รุ่นจึงสร้างกลุ่มที่เกี่ยวข้องกัน - สายพันธุ์จากนั้นจึงผสมพันธุ์และพันธุ์ที่มีคุณสมบัติทางพันธุกรรมที่มีลักษณะเฉพาะ
แม้ว่าการสังเกตและการเปรียบเทียบเหล่านี้ยังไม่อาจกลายเป็นพื้นฐานสำหรับการก่อตัวของวิทยาศาสตร์ แต่การพัฒนาอย่างรวดเร็วของการเลี้ยงสัตว์และการเพาะพันธุ์ เช่นเดียวกับการปลูกพืชและการผลิตเมล็ดพันธุ์ในช่วงครึ่งหลังของศตวรรษที่ 19 ทำให้เกิดความสนใจในการวิเคราะห์เพิ่มขึ้น ของปรากฏการณ์ทางพันธุกรรม
การพัฒนาวิทยาศาสตร์เกี่ยวกับการถ่ายทอดทางพันธุกรรมและความแปรปรวนได้รับการส่งเสริมอย่างมากโดยหลักคำสอนของชาร์ลส์ดาร์วินเกี่ยวกับต้นกำเนิดของสายพันธุ์ ซึ่งนำวิธีการทางประวัติศาสตร์ของการศึกษาวิวัฒนาการของสิ่งมีชีวิตมาสู่ชีววิทยา ดาร์วินเองก็ใช้ความพยายามอย่างมากในการศึกษาเรื่องพันธุกรรมและความแปรปรวน เขารวบรวมข้อเท็จจริงจำนวนมหาศาลและได้ข้อสรุปที่ถูกต้องหลายประการโดยอิงจากข้อเท็จจริงเหล่านั้น แต่เขาไม่สามารถกำหนดกฎแห่งกรรมพันธุ์ได้ ผู้ร่วมสมัยของเขาที่เรียกว่าลูกผสมซึ่งข้ามรูปแบบต่าง ๆ และมองหาระดับของความเหมือนและความแตกต่างระหว่างพ่อแม่และลูกหลานก็ไม่สามารถสร้างรูปแบบทั่วไปของการสืบทอดได้
เงื่อนไขอีกประการหนึ่งที่มีส่วนทำให้เกิดพันธุกรรมในฐานะวิทยาศาสตร์คือความก้าวหน้าในการศึกษาโครงสร้างและพฤติกรรมของเซลล์ร่างกายและเซลล์สืบพันธุ์ ย้อนกลับไปในทศวรรษที่ 70 ของศตวรรษที่ผ่านมา นักวิจัยทางเซลล์วิทยาจำนวนหนึ่ง (Chistyakov ในปี 1972, Strasburger ในปี 1875) ค้นพบการแบ่งเซลล์ร่างกายทางอ้อมที่เรียกว่า karyokinesis (Schleicher ในปี 1878) หรือไมโทซิส (Flemming ในปี 1882) . ในปี ค.ศ. 1888 ตามคำแนะนำของวัลไดรา องค์ประกอบถาวรของนิวเคลียสของเซลล์ถูกเรียกว่า "โครโมโซม" ในปีเดียวกันนั้น เฟลมมิงได้แบ่งวงจรการแบ่งเซลล์ทั้งหมดออกเป็นสี่ระยะหลัก ได้แก่ ระยะพยากรณ์ เมตาเฟส แอนาเฟส และเทโลเฟส
ควบคู่ไปกับการศึกษาการแบ่งเซลล์ร่างกาย ได้มีการวิจัยเกี่ยวกับการพัฒนาเซลล์สืบพันธุ์และกลไกการปฏิสนธิในสัตว์และพืช ในปี พ.ศ. 2419 O. Hertwig ได้ก่อตั้งเป็นครั้งแรกในเอไคโนเดิร์มซึ่งเป็นการรวมตัวของนิวเคลียสของอสุจิกับนิวเคลียสของไข่ เอ็น.เอ็น. Gorozhankin ในปี 1880 และ E. Strasburger ในปี 1884 ได้สร้างสิ่งเดียวกันสำหรับพืช: ครั้งแรก - สำหรับ gymnosperms, ที่สอง - สำหรับ angiosperms
ในปีเดียวกันนั้น Van Beneden (1883) และคนอื่นๆ เปิดเผยข้อเท็จจริงที่สำคัญว่าในระหว่างการพัฒนา เซลล์สืบพันธุ์ซึ่งแตกต่างจากเซลล์ร่างกาย ได้รับการลดจำนวนโครโมโซมลงครึ่งหนึ่งอย่างแน่นอน และในระหว่างการปฏิสนธิ - ฟิวชั่นของเพศหญิงและชาย นิวเคลียส - จำนวนโครโมโซมปกติจะถูกเรียกคืน ค่าคงที่สำหรับแต่ละประเภท ดังนั้นจึงแสดงให้เห็นว่าแต่ละสปีชีส์มีลักษณะเฉพาะด้วยโครโมโซมจำนวนหนึ่ง
ดังนั้นเงื่อนไขข้างต้นมีส่วนทำให้เกิดการเกิดขึ้นของพันธุศาสตร์ในฐานะวินัยทางชีววิทยาที่แยกจากกัน - วินัยที่มีวิชาและวิธีการวิจัยของตัวเอง
การกำเนิดทางพันธุศาสตร์อย่างเป็นทางการถือเป็นฤดูใบไม้ผลิของปี 1900 เมื่อนักพฤกษศาสตร์สามคนซึ่งเป็นอิสระจากกันในสามประเทศที่แตกต่างกันในสถานที่ต่าง ๆ ได้ค้นพบรูปแบบที่สำคัญที่สุดบางประการของการสืบทอดลักษณะในลูกหลาน ของลูกผสม G. de Vries (ฮอลแลนด์) จากการทำงานร่วมกับอีฟนิ่งพริมโรส ดอกป๊อปปี้ ดอก Datura และพืชอื่นๆ รายงานว่า "กฎของการแยกลูกผสม"; K. Correns (เยอรมนี) กำหนดรูปแบบของการแยกข้าวโพดและตีพิมพ์บทความ “กฎของ Gregor Mendel เกี่ยวกับพฤติกรรมของลูกหลานในเชื้อชาติลูกผสม”; ในปีเดียวกันนั้น K. Csermak (ออสเตรีย) ตีพิมพ์บทความ (เกี่ยวกับการข้ามเทียมใน Pisum Sativum)
วิทยาศาสตร์แทบไม่รู้จักการค้นพบที่ไม่คาดคิดเลย การค้นพบที่ยอดเยี่ยมที่สุดที่สร้างขั้นตอนในการพัฒนานั้นมักจะมีรุ่นก่อนอยู่เสมอ สิ่งนี้เกิดขึ้นพร้อมกับการค้นพบกฎแห่งกรรมพันธุ์ ปรากฎว่านักพฤกษศาสตร์สามคนที่ค้นพบรูปแบบของการแบ่งแยกในลูกหลานของลูกผสมที่มีลักษณะเฉพาะเพียง "ค้นพบ" รูปแบบของมรดกที่ค้นพบย้อนกลับไปในปี 1865 โดยเกรเกอร์ เมนเดล และสรุปโดยเขาในบทความ "การทดลองเกี่ยวกับพืชลูกผสม" ซึ่งตีพิมพ์ใน “การดำเนินการ” ของสมาคมนักวิทยาศาสตร์ธรรมชาติในบรุนน์ (เชโกสโลวาเกีย)
การใช้ต้นถั่ว G. Mendel ได้พัฒนาวิธีการวิเคราะห์ทางพันธุกรรมของการสืบทอดลักษณะเฉพาะของสิ่งมีชีวิตและสร้างปรากฏการณ์ที่สำคัญพื้นฐานสองประการ:
ลักษณะถูกกำหนดโดยปัจจัยทางพันธุกรรมส่วนบุคคลที่ถ่ายทอดผ่านเซลล์สืบพันธุ์
ลักษณะเฉพาะของสิ่งมีชีวิตจะไม่หายไประหว่างการผสมข้ามพันธุ์ แต่จะถูกเก็บรักษาไว้ในลูกหลานในรูปแบบเดียวกับที่อยู่ในสิ่งมีชีวิตต้นกำเนิด
สำหรับทฤษฎีวิวัฒนาการ หลักการเหล่านี้มีความสำคัญอย่างยิ่ง พวกเขาเปิดเผยแหล่งที่มาที่สำคัญที่สุดประการหนึ่งของความแปรปรวน กล่าวคือกลไกในการรักษาความเหมาะสมของคุณลักษณะของสายพันธุ์ต่างๆ มาหลายชั่วอายุคน หากลักษณะการปรับตัวของสิ่งมีชีวิตที่เกิดขึ้นภายใต้การควบคุมของการคัดเลือกถูกดูดซับและหายไประหว่างการผสมข้ามพันธุ์ ความก้าวหน้าของสายพันธุ์ก็จะเป็นไปไม่ได้
การพัฒนาทางพันธุศาสตร์ในเวลาต่อมาทั้งหมดเกี่ยวข้องกับการศึกษาและการขยายหลักการเหล่านี้และการประยุกต์กับทฤษฎีวิวัฒนาการและการคัดเลือก
จากหลักการพื้นฐานที่กำหนดไว้ของ Mendel ปัญหาทั้งหมดตามมาอย่างมีเหตุผล ซึ่งทีละขั้นตอนจะได้รับวิธีแก้ปัญหาเมื่อพันธุกรรมพัฒนาขึ้น ในปี 1901 เดอ ไวรีส์ได้กำหนดทฤษฎีการกลายพันธุ์ ซึ่งระบุว่าคุณสมบัติและลักษณะทางพันธุกรรมของสิ่งมีชีวิตเปลี่ยนแปลงอย่างกะทันหัน - กลายพันธุ์
ในปี 1903 นักสรีรวิทยาพืชชาวเดนมาร์ก V. Johannsen ตีพิมพ์ผลงานเรื่อง "On Inheritance in Populations and Pure Lines" ซึ่งมีการทดลองแล้วว่าพืชที่คล้ายกันภายนอกซึ่งอยู่ในพันธุ์เดียวกันนั้นมีความแตกต่างทางพันธุกรรม - พวกมันประกอบเป็นประชากร ประชากรประกอบด้วยบุคคลที่แตกต่างกันทางพันธุกรรมหรือกลุ่มที่เกี่ยวข้องกัน - เส้น ในการศึกษาเดียวกัน แสดงให้เห็นอย่างชัดเจนที่สุดว่ามีความแปรปรวนในสิ่งมีชีวิตสองประเภท: กรรมพันธุ์ กำหนดโดยยีน และไม่ใช่ทางพันธุกรรม กำหนดโดยการสุ่มผสมของปัจจัยที่ทำหน้าที่ในการสำแดงลักษณะ
ในขั้นต่อไปของการพัฒนาพันธุศาสตร์ได้รับการพิสูจน์แล้วว่ารูปแบบทางพันธุกรรมมีความเกี่ยวข้องกับโครโมโซม ข้อเท็จจริงประการแรกที่เปิดเผยบทบาทของโครโมโซมในการถ่ายทอดทางพันธุกรรมคือการพิสูจน์บทบาทของโครโมโซมในการกำหนดเพศในสัตว์และการค้นพบกลไกการแยกเพศแบบ 1:1
ตั้งแต่ปี พ.ศ. 2454 ที. มอร์แกนและเพื่อนร่วมงานของเขาที่มหาวิทยาลัยโคลัมเบียในสหรัฐอเมริกาเริ่มตีพิมพ์ผลงานหลายชุดซึ่งเขาได้กำหนดทฤษฎีพันธุกรรมของโครโมโซม จากการทดลองพิสูจน์ว่าพาหะหลักของยีนคือโครโมโซม และยีนนั้นอยู่บนโครโมโซมเป็นเส้นตรง
ในปี พ.ศ. 2465 N.I. วาวิลอฟกำหนดกฎของอนุกรมความคล้ายคลึงกันในความแปรปรวนทางพันธุกรรม ตามที่ชนิดพันธุ์พืชและสัตว์ที่เกี่ยวข้องโดยแหล่งกำเนิดมีอนุกรมความแปรปรวนทางพันธุกรรมที่คล้ายคลึงกัน การใช้กฎหมายนี้ N.I. Vavilov ได้ก่อตั้งศูนย์กลางของต้นกำเนิดของพืชที่ปลูกซึ่งมีรูปแบบทางพันธุกรรมที่หลากหลายมากที่สุด
ในปี 1925 ในประเทศของเรา G.A. แนดสัน และ G.S. Filippov เกี่ยวกับเห็ดและในปี 1927 G. Möller ในสหรัฐอเมริกาเกี่ยวกับแมลงวันผลไม้ Drosophila ได้รับหลักฐานเกี่ยวกับอิทธิพลของรังสีเอกซ์ต่อการเกิดการเปลี่ยนแปลงทางพันธุกรรม ในขณะเดียวกันก็แสดงให้เห็นว่าอัตราการกลายพันธุ์เพิ่มขึ้นมากกว่า 100 เท่า การศึกษาเหล่านี้พิสูจน์ความแปรปรวนของยีนภายใต้อิทธิพลของปัจจัยด้านสิ่งแวดล้อม การพิสูจน์อิทธิพลของรังสีไอออไนซ์ต่อการเกิดการกลายพันธุ์นำไปสู่การสร้างสาขาใหม่ของพันธุศาสตร์ - พันธุศาสตร์การแผ่รังสีซึ่งมีความสำคัญมากยิ่งขึ้นด้วยการค้นพบพลังงานปรมาณู
ในปีพ. ศ. 2477 T. Paynter โดยใช้โครโมโซมขนาดยักษ์ของต่อมน้ำลายของ Dipterans พิสูจน์ว่าความไม่ต่อเนื่องของโครงสร้างทางสัณฐานวิทยาของโครโมโซมซึ่งแสดงออกมาในรูปแบบของดิสก์ต่าง ๆ สอดคล้องกับตำแหน่งของยีนในโครโมโซมซึ่งสร้างขึ้นก่อนหน้านี้โดยพันธุกรรมล้วนๆ วิธีการ การค้นพบครั้งนี้ถือเป็นจุดเริ่มต้นของการศึกษาโครงสร้างและการทำงานของยีนในเซลล์
ในช่วงทศวรรษที่ 40 จนถึงปัจจุบัน มีการค้นพบปรากฏการณ์ทางพันธุกรรมใหม่ทั้งหมด (โดยหลักคือจุลินทรีย์) ซึ่งเผยให้เห็นความเป็นไปได้ในการวิเคราะห์โครงสร้างยีนในระดับโมเลกุล ในช่วงไม่กี่ปีที่ผ่านมา ด้วยการแนะนำวิธีการวิจัยใหม่ๆ ในด้านพันธุศาสตร์ ซึ่งยืมมาจากจุลชีววิทยา เราได้ค้นพบวิธีแก้ปัญหาว่ายีนควบคุมลำดับของกรดอะมิโนในโมเลกุลโปรตีนอย่างไร
ประการแรกควรกล่าวว่าขณะนี้ได้รับการพิสูจน์แล้วอย่างสมบูรณ์ว่าพาหะของพันธุกรรมคือโครโมโซมซึ่งประกอบด้วยกลุ่มโมเลกุลดีเอ็นเอ
ทำการทดลองที่ค่อนข้างง่าย: DNA บริสุทธิ์ถูกแยกออกจากแบคทีเรียที่ถูกฆ่าในสายพันธุ์หนึ่งที่มีลักษณะภายนอกพิเศษและถ่ายโอนไปยังแบคทีเรียที่มีชีวิตในสายพันธุ์อื่น หลังจากนั้นแบคทีเรียที่สืบพันธุ์ของสายพันธุ์หลังจะได้ลักษณะของสายพันธุ์แรก การทดลองที่คล้ายกันจำนวนมากแสดงให้เห็นว่า DNA เป็นพาหะของพันธุกรรม
ในปี 1953 F. Crick (อังกฤษ) และ J. Watstone (สหรัฐอเมริกา) ถอดรหัสโครงสร้างของโมเลกุล DNA พวกเขาพบว่าโมเลกุล DNA แต่ละโมเลกุลประกอบด้วยสายพอลิดีออกซีไรโบนิวคลีอิกสองเส้นที่บิดเกลียวเป็นเกลียวรอบแกนร่วม
ปัจจุบันพบแนวทางในการแก้ปัญหาการจัดระเบียบรหัสพันธุกรรมและถอดรหัสการทดลอง พันธุศาสตร์ พร้อมด้วยชีวเคมีและชีวฟิสิกส์ เกือบจะช่วยอธิบายกระบวนการสังเคราะห์โปรตีนในเซลล์และการสังเคราะห์โมเลกุลโปรตีนโดยสังเคราะห์แล้ว สิ่งนี้เป็นการเริ่มต้นขั้นตอนใหม่อย่างสมบูรณ์ในการพัฒนาไม่เพียงแต่พันธุกรรมเท่านั้น แต่ยังรวมถึงชีววิทยาทั้งหมดโดยรวมด้วย
การพัฒนาทางพันธุศาสตร์มาจนถึงทุกวันนี้เป็นรากฐานของการวิจัยที่เพิ่มขึ้นอย่างต่อเนื่องเกี่ยวกับความแตกต่างในการทำงาน สัณฐานวิทยา และชีวเคมีของโครโมโซม มีการดำเนินการหลายอย่างในพื้นที่นี้ มีการดำเนินการหลายอย่างแล้ว และทุกๆ วัน วิทยาศาสตร์ที่ล้ำหน้ากำลังเข้าใกล้เป้าหมาย - เผยให้เห็นธรรมชาติของยีน จนถึงปัจจุบัน มีการสร้างปรากฏการณ์หลายประการที่แสดงถึงลักษณะของยีน ประการแรก ยีนบนโครโมโซมมีคุณสมบัติในการสืบพันธุ์ด้วยตนเอง (การผลิตอัตโนมัติ) ประการที่สอง มันสามารถเปลี่ยนแปลงการกลายพันธุ์ได้ ประการที่สามมีความเกี่ยวข้องกับโครงสร้างทางเคมีบางอย่างของกรดดีออกซีไรโบนิวคลีอิก - DNA; ประการที่สี่ ควบคุมการสังเคราะห์กรดอะมิโนและลำดับในโมเลกุลโปรตีน ในการเชื่อมต่อกับการวิจัยล่าสุด แนวคิดใหม่เกี่ยวกับยีนในฐานะระบบการทำงานกำลังถูกสร้างขึ้น และผลกระทบของยีนต่อการกำหนดลักษณะจะได้รับการพิจารณาในระบบบูรณาการของยีน - จีโนไทป์
แนวโน้มใหม่ของการสังเคราะห์สิ่งมีชีวิตดึงดูดความสนใจอย่างมากจากนักพันธุศาสตร์ นักชีวเคมี นักฟิสิกส์ และผู้เชี่ยวชาญอื่นๆ
>บทคัดย่อทางชีววิทยา
พันธุศาสตร์เป็นหนึ่งในสาขาที่สำคัญที่สุดของชีววิทยา นี่คือศาสตร์แห่งกฎแห่งกรรมพันธุ์และความแปรปรวน คำว่า "พันธุศาสตร์" มีต้นกำเนิดจากภาษากรีกและแปลว่า "มาจากใครบางคน" วัตถุประสงค์ของการวิจัยอาจเป็นพืช สัตว์ คน จุลินทรีย์ พันธุศาสตร์มีความเกี่ยวข้องอย่างใกล้ชิดกับวิทยาศาสตร์ เช่น พันธุวิศวกรรม การแพทย์ จุลชีววิทยา และอื่นๆ
ในขั้นต้น พันธุกรรมถือเป็นรูปแบบของพันธุกรรมและความแปรปรวนโดยพิจารณาจากลักษณะภายนอกและภายในของสิ่งมีชีวิต ปัจจุบัน เป็นที่ทราบกันดีว่ามียีนอยู่และมีส่วนพิเศษของ DNA หรือ RNA ซึ่งก็คือโมเลกุลที่ใช้ตั้งโปรแกรมข้อมูลทางพันธุกรรมทั้งหมด
จากหลักฐานทางโบราณคดี ผู้คนทราบมานานกว่า 6,000 ปีแล้วว่าลักษณะทางกายภาพบางอย่างสามารถสืบทอดจากรุ่นสู่รุ่นได้ มนุษย์ยังได้เรียนรู้ที่จะสร้างพันธุ์พืชและพันธุ์สัตว์ที่ได้รับการปรับปรุงโดยการเลือกประชากรบางกลุ่มและผสมข้ามพันธุ์กัน อย่างไรก็ตาม ความสำคัญของพันธุศาสตร์กลายเป็นที่รู้จักอย่างสมบูรณ์เฉพาะในศตวรรษที่ 19 และ 20 เท่านั้นที่มีการถือกำเนิดของกล้องจุลทรรศน์สมัยใหม่ พระภิกษุชาวออสเตรีย Gregor Mendel มีส่วนช่วยอย่างมากในการพัฒนาพันธุศาสตร์ ในปีพ.ศ. 2409 เขาได้นำเสนอผลงานของเขาเกี่ยวกับรากฐานของพันธุศาสตร์สมัยใหม่ เขาพิสูจน์ว่าความโน้มเอียงทางพันธุกรรมไม่ได้ปะปนกัน แต่ส่งต่อจากรุ่นสู่รุ่นในรูปแบบของหน่วยที่แยกจากกัน ในปี 1912 โทมัส มอร์แกน นักพันธุศาสตร์ชาวอเมริกันได้พิสูจน์ว่าหน่วยเหล่านี้อยู่ในโครโมโซม ตั้งแต่นั้นเป็นต้นมา พันธุศาสตร์คลาสสิกได้ก้าวไปข้างหน้าทางวิทยาศาสตร์และมีความก้าวหน้าอย่างมากในการอธิบายพันธุกรรมไม่เพียงแต่ในระดับสิ่งมีชีวิตเท่านั้น แต่ยังรวมถึงระดับยีนด้วย
ยุคของอณูพันธุศาสตร์เริ่มขึ้นในทศวรรษที่ 1940 และ 1950 มีหลักฐานแสดงบทบาทนำของ DNA ในการถ่ายทอดข้อมูลทางพันธุกรรม การค้นพบนี้คือการถอดรหัสโครงสร้าง DNA รหัสแฝด และการอธิบายกลไกของการสังเคราะห์โปรตีน นอกจากนี้ยังค้นพบลำดับกรดอะมิโนหรือนิวคลีโอไทด์ของ DNA และ RNA
การทดลองครั้งแรกในรัสเซียเกิดขึ้นในศตวรรษที่ 18 และเกี่ยวข้องกับการผสมพันธุ์พืช ในศตวรรษที่ 20 งานสำคัญปรากฏในพฤกษศาสตร์ทดลองและสัตววิทยา รวมถึงที่สถานีเกษตรทดลอง ในช่วงปลายทศวรรษที่ 1930 เครือข่ายสถาบันวิจัย สถานีทดลอง และแผนกพันธุศาสตร์ของมหาวิทยาลัยปรากฏขึ้นในประเทศ ในปี 1948 พันธุศาสตร์ได้รับการประกาศให้เป็นวิทยาศาสตร์เทียม การฟื้นฟูวิทยาศาสตร์เกิดขึ้นหลังจากการค้นพบและถอดรหัสโครงสร้างของ DNA ประมาณทศวรรษ 1960
หลายๆ คนเชื่อว่าสาขาพันธุศาสตร์ที่น่าสนใจที่สุดคือ พันธุศาสตร์ของมนุษย์– ศาสตร์แห่งการถ่ายทอดทางพันธุกรรมและความแปรปรวนของลักษณะนิสัยในมนุษย์ แท้จริงแล้ว ในพื้นที่นี้เองที่มีการอภิปรายทางวิทยาศาสตร์อย่างเผ็ดร้อนเกิดขึ้น และเป็นที่ที่ใช้วิธีการและเทคโนโลยีทางวิทยาศาสตร์ที่ทันสมัยที่สุด
มนุษย์อยู่ภายใต้กฎหมายมรดกเช่นเดียวกับสัตว์ใดๆ ที่มีวิธีการสืบพันธุ์แบบอาศัยเพศ เครื่องมือทางพันธุกรรมของมนุษย์นั้นเหมือนกับเครื่องมือของผู้อาศัยอื่น ๆ ในโลก พื้นฐานของมันคือ DNA ซึ่ง RNA ถูกสังเคราะห์ซึ่งจะทำหน้าที่ในการสังเคราะห์โปรตีน ความหลากหลายของยีนทั้งหมดถูกสร้างขึ้นด้วยการมีส่วนร่วมของนิวคลีโอไทด์สี่ตัว ข้อมูลทางพันธุกรรมถูกอ่านเป็นแฝดสาม ยิ่งไปกว่านั้น ยีนบางชนิดในสิ่งมีชีวิตชนิดที่ไม่เกี่ยวข้องกันโดยสิ้นเชิงนั้นเหมือนกันโดยสิ้นเชิง เป็นเรื่องยากที่จะจินตนาการ แต่มนุษย์และกล้วยมียีนโครงสร้างครึ่งหนึ่งของยีนทั้งหมดเหมือนกัน! และความคล้ายคลึงกันระหว่างมนุษย์กับลิงชิมแปนซีคือ 98.7% ของยีน ยิ่งไปกว่านั้น ไม่เพียงแต่ยีนที่ทำงานตามปกติจะเหมือนกันเท่านั้น แต่ยังเหมือนกันอีกด้วย เทียม - ส่วนของโครโมโซมที่คล้ายกับยีนโครงสร้าง แต่มี "ข้อผิดพลาดในการพิมพ์" ที่ทำให้โครโมโซมไม่ทำงาน
มีการเปรียบเทียบอันชาญฉลาดประการหนึ่งเกี่ยวกับความคล้ายคลึงทางพันธุกรรมของสิ่งมีชีวิตทุกชนิด ลองนึกภาพว่าเพื่อนร่วมโต๊ะสองคนเขียนเรียงความซึ่งไม่เพียงแต่เนื้อหาจะเหมือนกัน แต่ยังรวมถึงข้อผิดพลาดด้วย บทความในหน้าสมุดบันทึกสามหน้าแตกต่างกันเพียงคำเดียวในข้อความ เห็นได้ชัดว่านักเรียนคัดลอกเรียงความจากกันหรือจากหนังสือเล่มเดียวกัน นี่คือความคล้ายคลึงกันอย่างมากในโครงสร้างของ "ศักดิ์สิทธิ์แห่งความศักดิ์สิทธิ์" - เครื่องมือแห่งพันธุกรรม - ซึ่งเป็นข้อพิสูจน์ที่หักล้างไม่ได้ถึงความเป็นเอกภาพแห่งต้นกำเนิดของทุกสิ่ง ชีวิตบนโลกของเรา
การถ่ายทอดลักษณะในมนุษย์ขึ้นอยู่กับกฎหมายและกฎเกณฑ์ทางพันธุศาสตร์: กฎของเมนเดล, มอร์แกน, การเชื่อมโยงของยีน, ปฏิสัมพันธ์ของยีนอัลลีลิกและไม่ใช่อัลลีลิก (ตารางที่ 1, 2) อย่างไรก็ตาม เนื่องจากบุคคลไม่เพียงแต่เป็นสิ่งมีชีวิตทางชีววิทยาเท่านั้น แต่ยังเป็นสิ่งมีชีวิตทางสังคมด้วย การศึกษาทางพันธุกรรมของสายพันธุ์ดังกล่าว โฮโม เซเปียนส์ แตกต่างกันหลายประการ ดังนี้
ตารางที่ 1. ลักษณะเด่นและลักษณะด้อยบางประการในร่างกายมนุษย์
| เข้าสู่ระบบ | ที่เด่น | ถอย |
| ขนาดตา | ใหญ่ | เด็กน้อย |
| สีตา | สีน้ำตาล | สีฟ้า |
| ประเภทตา | มองโกลอยด์ | คนผิวขาว |
| รูปร่างตา | ตรง | เป๋ |
| วิสัยทัศน์ | สายตาสั้น | บรรทัดฐาน |
| ตาเหล่ | กิน | เลขที่ |
| คาง | กว้าง | เผ็ด |
| คาง | ยาว | สั้น |
| คางกลวง | กิน | เลขที่ |
| รูปร่างริมฝีปาก | กว้าง | บาง |
| กระ | กิน | เลขที่ |
| ความหนาแน่นของคิ้ว | กว้างหนา | แคบหายาก |
| รูปร่างคิ้ว | เชื่อมต่อที่ดั้งจมูก | อย่าเชื่อมบนดั้งจมูก |
| ขนตา | ยาว | สั้น |
| รูปร่างจมูก | กลม | เผ็ด |
| รูปร่างจมูก | ตรง | ดูแคลน |
| ขนาดจมูก | ใหญ่ | ปกติ |
| สะพานจมูก | มีโคก | ตรง |
| ปลายผมบนศีรษะ | กิน | เลขที่ |
| โครงสร้างเส้นผม | แข็ง | อ่อนนุ่ม |
| สีผม | มืด | แสงสว่าง |
| เสียงผู้หญิง | โซปราโน | อัลโต |
| เสียงผู้ชาย | เบส | เทเนอร์ |
| ความสูง | สั้น | สูง |
| จำนวนนิ้ว | โพลีฟิงเกอร์ | ห้านิ้ว |
| กรุ๊ปเลือด | II, III, IV | 1 |
| ปัจจัย Rh เลือด | เชิงบวก | เชิงลบ |
| สีผิว | มืด | แสงสว่าง |
| โครงสร้างผิวหนัง | อ้วน | บาง |
| มือนำ | ซ้าย | ขวา |
| รูปหน้า | กลม | เป็นรูปสี่เหลี่ยมผืนผ้า |
ตารางที่ 2. ลักษณะที่มีการครอบงำที่ไม่สมบูรณ์ในมนุษย์วัสดุจากเว็บไซต์
| เข้าสู่ระบบ | ที่เด่นฟีโนไทป์ | ฟีโนไทป์เฮเทอโรไซกัส | ถอยฟีโนไทป์ |
| ขนาดจมูก | ใหญ่ | เฉลี่ย | เล็ก |
| ขนาดปาก | ใหญ่ | เฉลี่ย | เล็ก |
| ขนาดตา | ใหญ่ | เฉลี่ย | เด็กน้อย |
| ระยะห่างระหว่างดวงตา | ใหญ่ | เฉลี่ย | เล็ก |
| สีคิ้ว | มืดมาก | มืด | แสงสว่าง |
| ผมหยิก | หยิกงอ | หยิกงอ | โดยตรง |
พันธุศาสตร์มนุษย์ — ศาสตร์แห่งการถ่ายทอดทางพันธุกรรมและความแปรปรวนของลักษณะในมนุษย์ การศึกษาจีโนมมนุษย์ได้ยืนยันต้นกำเนิดวิวัฒนาการของมันอย่างไม่คลุมเครือ ในพันธุศาสตร์มนุษย์ เป็นไปไม่ได้เลยที่จะใช้วิธีการทางพันธุกรรมแบบดั้งเดิมบางวิธี โดยเฉพาะวิธีการผสมพันธุ์ ปัจจัยทางสังคมยังทิ้งร่องรอยไว้ที่กระบวนการทางพันธุกรรมที่เกิดขึ้นในประชากรมนุษย์
พันธุศาสตร์การแพทย์ - ศาสตร์แห่งปัญหาทางการแพทย์ด้านพันธุกรรม มีโรคทางพันธุกรรมและโรคที่มีความบกพร่องทางพันธุกรรม โรคทางพันธุกรรมแบ่งออกเป็นโรคทางพันธุกรรมโครโมโซมและจีโนม
ในหน้านี้จะมีเนื้อหาในหัวข้อต่อไปนี้:
คำถามเกี่ยวกับเนื้อหานี้:
แม้ว่าประวัติศาสตร์ด้านพันธุศาสตร์จะเริ่มขึ้นในศตวรรษที่ 19 แม้แต่คนโบราณก็ยังสังเกตเห็นว่าสัตว์และพืชถ่ายทอดลักษณะเฉพาะของตนมาหลายชั่วอายุคน กล่าวอีกนัยหนึ่งก็ชัดเจนว่าพันธุกรรมนั้นมีอยู่ในธรรมชาติ ในกรณีนี้ คุณลักษณะส่วนบุคคลอาจมีการเปลี่ยนแปลง นั่นคือนอกเหนือจากพันธุกรรมแล้วยังมีความแปรปรวนในธรรมชาติอีกด้วย พันธุกรรมและความแปรปรวนเป็นคุณสมบัติพื้นฐานของสิ่งมีชีวิต เป็นเวลานาน (จนถึงศตวรรษที่ 19-20) เหตุผลที่แท้จริงของการดำรงอยู่ของพวกเขาถูกซ่อนไม่ให้ผู้คนเห็น สิ่งนี้ทำให้เกิดสมมติฐานจำนวนหนึ่งที่สามารถแบ่งออกเป็นสองประเภท: มรดกทางตรงและมรดกทางอ้อม
ผู้ติดตาม มรดกโดยตรง(ฮิปโปเครติส ลามาร์ก ดาร์วิน ฯลฯ) สันนิษฐานว่าข้อมูลจากแต่ละอวัยวะและแต่ละส่วนของร่างกายของสิ่งมีชีวิตต้นกำเนิดถูกส่งไปยังสิ่งมีชีวิตรุ่นลูกผ่านสารบางชนิด (เจมูลีสตามดาร์วิน) ซึ่งรวบรวมไว้ในผลิตภัณฑ์สืบพันธุ์ จากข้อมูลของลามาร์ก ความเสียหายหรือการพัฒนาที่แข็งแกร่งของอวัยวะจะถูกส่งไปยังคนรุ่นต่อไปโดยตรง สมมติฐาน มรดกทางอ้อม(อริสโตเติลในศตวรรษที่ 4 ก่อนคริสตกาล ไวส์แมนในศตวรรษที่ 19) แย้งว่าผลิตภัณฑ์สืบพันธุ์ถูกสร้างขึ้นแยกจากกันในร่างกายและ "ไม่รู้" เกี่ยวกับการเปลี่ยนแปลงในอวัยวะต่างๆ ของร่างกาย
ไม่ว่าในกรณีใด สมมติฐานทั้งสองต่างแสวงหา "รากฐาน" ของพันธุกรรมและการแปรผัน
ประวัติความเป็นมาของพันธุศาสตร์ในฐานะวิทยาศาสตร์เริ่มต้นด้วยงานของ Gregor Mendel (1822-1884) ซึ่งในยุค 60 ได้ทำการทดลองอย่างเป็นระบบและมากมายเกี่ยวกับถั่ว ได้สร้างรูปแบบการถ่ายทอดทางพันธุกรรมจำนวนหนึ่ง และได้ตั้งสมมติฐานเกี่ยวกับการจัดระเบียบของสารพันธุกรรมเป็นครั้งแรก . การเลือกวัตถุประสงค์การศึกษาที่ถูกต้อง ลักษณะที่กำลังศึกษา รวมถึงโชคทางวิทยาศาสตร์ทำให้เขาสามารถกำหนดกฎสามข้อได้:
เมนเดลตระหนักว่าเนื้อหาทางพันธุกรรมนั้นแยกจากกัน แสดงให้เห็นโดยความโน้มเอียงส่วนบุคคลที่ส่งต่อไปยังลูกหลาน นอกจากนี้ความโน้มเอียงแต่ละอย่างยังมีส่วนรับผิดชอบต่อการพัฒนาลักษณะเฉพาะของสิ่งมีชีวิต ลักษณะนี้เกิดจากความโน้มเอียงคู่หนึ่งที่มาพร้อมกับเซลล์สืบพันธุ์จากทั้งพ่อและแม่
ในเวลานั้น การค้นพบทางวิทยาศาสตร์ของ Mendel ไม่ได้ให้ความสำคัญมากนัก กฎของมันถูกค้นพบอีกครั้งเมื่อต้นศตวรรษที่ 20 โดยนักวิทยาศาสตร์หลายคนโดยใช้พืชและสัตว์ต่างกัน
ในช่วงทศวรรษที่ 80 ของศตวรรษที่ 19 มีการอธิบายไมโทซิสและไมโอซิส ในระหว่างที่มีการกระจายโครโมโซมระหว่างเซลล์ลูกสาวเป็นประจำ ในตอนต้นของศตวรรษที่ 20 T. Boveri และ W. Setton ได้ข้อสรุปว่า ความต่อเนื่องของคุณสมบัติในสิ่งมีชีวิตหลายชั่วอายุคนนั้นพิจารณาจากความต่อเนื่องของโครโมโซม- นั่นคือในช่วงเวลานี้โลกวิทยาศาสตร์เข้าใจในโครงสร้างของ "สารตั้งต้น" ของการถ่ายทอดทางพันธุกรรม
ดับเบิลยู. แบทสันถูกค้นพบ กฎแห่งความบริสุทธิ์ของ gameteและศาสตร์แห่งพันธุกรรมและความแปรปรวนถูกเรียกโดยเขาเป็นครั้งแรกในประวัติศาสตร์ พันธุศาสตร์- V. Johansen ได้นำแนวความคิดนี้มาสู่วิทยาศาสตร์ (พ.ศ. 2452) จีโนไทป์และฟีโนไทป์- ในเวลานั้นนักวิทยาศาสตร์ได้ตระหนักรู้แล้วว่า ยีนเป็นปัจจัยทางพันธุกรรมเบื้องต้น- แต่ยังไม่ทราบลักษณะทางเคมีของมัน
ในปีพ.ศ. 2449 ได้มีการเปิดทำการ ปรากฏการณ์การเชื่อมโยงของยีน, รวมทั้ง การถ่ายทอดลักษณะทางเพศที่เชื่อมโยง- แนวคิดเรื่องจีโนไทป์เน้นย้ำว่ายีนของสิ่งมีชีวิตไม่ได้เป็นเพียงชุดของหน่วยพันธุกรรมที่เป็นอิสระเท่านั้น แต่ยังสร้างระบบที่สังเกตการพึ่งพาบางอย่างด้วย
ควบคู่ไปกับการศึกษาเรื่องพันธุกรรม การค้นพบรูปแบบของความแปรปรวนก็เกิดขึ้น ในปี 1901 เดอ ไวรีส์ได้วางรากฐานสำหรับหลักคำสอนเรื่องความแปรปรวนของการกลายพันธุ์ที่เกี่ยวข้องกับการเกิดการเปลี่ยนแปลงของโครโมโซม ซึ่งนำไปสู่การเปลี่ยนแปลงในลักษณะต่างๆ หลังจากนั้นไม่นานก็พบว่าโครโมโซมมักเกิดขึ้นเมื่อสัมผัสกับรังสี สารเคมีบางชนิด ฯลฯ ดังนั้นจึงได้รับการพิสูจน์แล้วว่าโครโมโซมไม่เพียงเป็น "สารตั้งต้น" ของการถ่ายทอดทางพันธุกรรมเท่านั้น แต่ยังมีความแปรปรวนอีกด้วย
ในปี 1910 กลุ่มของที. มอร์แกนได้พัฒนาขึ้นโดยสรุปการค้นพบก่อนหน้านี้เป็นส่วนใหญ่ ทฤษฎีโครโมโซม:
ยีนอยู่บนโครโมโซมและจัดเรียงเป็นเส้นตรงตรงนั้น
โครโมโซมแต่ละตัวมีโครโมโซมคล้ายคลึงกัน
ลูกจะได้รับโครโมโซมคล้ายคลึงกันจากพ่อแม่แต่ละคน
โครโมโซมที่คล้ายคลึงกันประกอบด้วยยีนชุดเดียวกัน แต่อัลลีลของยีนอาจแตกต่างกัน
ยีนที่อยู่บนโครโมโซมเดียวกันจะสืบทอดร่วมกัน() หากอยู่ใกล้กัน
เหนือสิ่งอื่นใด extrachromosomal หรือ cytoplasmic มีการค้นพบมรดกที่เกี่ยวข้องกับไมโตคอนเดรียและคลอโรพลาสต์เมื่อต้นศตวรรษที่ 20
การวิเคราะห์ทางเคมีของโครโมโซมแสดงให้เห็นว่าประกอบด้วยโปรตีนและกรดนิวคลีอิก ในช่วงครึ่งแรกของศตวรรษที่ 20 นักวิทยาศาสตร์หลายคนมีแนวโน้มที่จะเชื่อว่าโปรตีนเป็นพาหะของพันธุกรรมและความแปรปรวน
ในช่วงทศวรรษที่ 40 ของศตวรรษที่ 20 การก้าวกระโดดเกิดขึ้นในประวัติศาสตร์ของพันธุศาสตร์ การวิจัยกำลังก้าวไปสู่ระดับโมเลกุล
ในปีพ.ศ. 2487 ได้มีการค้นพบว่าสารในเซลล์ เช่น มีหน้าที่รับผิดชอบต่อลักษณะทางพันธุกรรม DNA ได้รับการยอมรับว่าเป็นพาหะของข้อมูลทางพันธุกรรมต่อมาก็กำหนดไว้ว่า ยีนหนึ่งเข้ารหัสโพลีเปปไทด์หนึ่งตัว.
ในปี 1953 D. Watson และ F. Crick ได้ถอดรหัสโครงสร้างของ DNA ปรากฎว่าสิ่งนี้ เกลียวคู่ที่ประกอบด้วยนิวคลีโอไทด์- พวกเขาสร้างแบบจำลองเชิงพื้นที่ของโมเลกุลดีเอ็นเอ
ต่อมามีการค้นพบคุณสมบัติต่อไปนี้ (ยุค 60):
กรดอะมิโนแต่ละตัวของโพลีเปปไทด์ถูกเข้ารหัสโดยแฝด(เบสไนโตรเจนสามเบสใน DNA)
กรดอะมิโนแต่ละตัวถูกเข้ารหัสโดยแฝดตั้งแต่หนึ่งตัวขึ้นไป
แฝดสามไม่ทับซ้อนกัน
การอ่านเริ่มต้นด้วยแฝดสามเริ่มต้น
ไม่มี "เครื่องหมายวรรคตอน" ใน DNA
ในยุค 70 การก้าวกระโดดเชิงคุณภาพอีกครั้งเกิดขึ้นในประวัติศาสตร์ของพันธุศาสตร์ - การพัฒนา พันธุวิศวกรรม- นักวิทยาศาสตร์เริ่มต้น สังเคราะห์ยีน เปลี่ยนจีโนม- ในเวลานี้พวกเขากำลังศึกษาอย่างกระตือรือร้น กลไกระดับโมเลกุลที่เป็นรากฐานของกระบวนการทางสรีรวิทยาต่างๆ.
ในยุค 90 จีโนมถูกเรียงลำดับ(ถอดรหัสลำดับนิวคลีโอไทด์ใน DNA) ของสิ่งมีชีวิตหลายชนิด ในปี พ.ศ. 2546 โครงการหาลำดับจีโนมมนุษย์เสร็จสมบูรณ์ ปัจจุบันก็มี ฐานข้อมูลจีโนม- ทำให้สามารถศึกษาลักษณะทางสรีรวิทยา โรคของมนุษย์และสิ่งมีชีวิตอื่น ๆ ได้อย่างครอบคลุม รวมถึงระบุความสัมพันธ์ระหว่างสายพันธุ์ด้วย อย่างหลังอนุญาตให้อนุกรมวิธานของสิ่งมีชีวิตก้าวไปสู่ระดับใหม่
การกำเนิดของพันธุกรรมในช่วงเปลี่ยนผ่านของสองศตวรรษ (พ.ศ. 2443) ได้รับการจัดเตรียมโดยการพัฒนาทางวิทยาศาสตร์ชีวภาพก่อนหน้านี้ทั้งหมด ศตวรรษที่สิบเก้า เข้าสู่ประวัติศาสตร์ชีววิทยาด้วยการค้นพบที่ยิ่งใหญ่สองประการ ได้แก่ ทฤษฎีเซลล์ที่คิดค้นโดย M. Schleiden และ T. Schwann (1838) และหลักคำสอนเชิงวิวัฒนาการของ Charles Darwin (1859) การค้นพบทั้งสองมีบทบาทสำคัญในการพัฒนาพันธุศาสตร์ ทฤษฎีเซลล์ซึ่งประกาศว่าเซลล์เป็นหน่วยโครงสร้างพื้นฐานและหน้าที่ของสิ่งมีชีวิตทุกชนิด กระตุ้นให้เกิดความสนใจมากขึ้นในการศึกษาโครงสร้างของมัน ซึ่งต่อมานำไปสู่การค้นพบโครโมโซมและคำอธิบายกระบวนการแบ่งเซลล์ ในทางกลับกันทฤษฎีของชาร์ลส์ดาร์วินเกี่ยวข้องกับคุณสมบัติที่สำคัญที่สุดของสิ่งมีชีวิตซึ่งต่อมาได้กลายเป็นหัวข้อของการศึกษาทางพันธุกรรม - พันธุกรรมและความแปรปรวน ทั้งสองทฤษฎีเมื่อปลายศตวรรษที่ 19 รวมเข้าด้วยกันโดยแนวคิดถึงความจำเป็นในการมีอยู่ของตัวพาวัสดุของคุณสมบัติเหล่านี้ซึ่งควรอยู่ในเซลล์
จนกระทั่งต้นศตวรรษที่ยี่สิบ สมมติฐานทั้งหมดเกี่ยวกับกลไกของการถ่ายทอดทางพันธุกรรมเป็นเพียงการเก็งกำไรเท่านั้น ดังนั้น ตามทฤษฎีการกำเนิดของตับโดย Charles Darwin (1868) อนุภาคเล็กๆ จะถูกแยกออกจากเซลล์ทั้งหมดของร่างกาย - gemmules ซึ่งไหลเวียนผ่านกระแสเลือดและเข้าสู่เซลล์สืบพันธุ์ หลังจากการหลอมรวมของเซลล์สืบพันธุ์ ในระหว่างการพัฒนาสิ่งมีชีวิตใหม่ เซลล์ประเภทเดียวกันที่มันถูกสร้างขึ้นจากแต่ละ gemmule มีคุณสมบัติทั้งหมด รวมถึงคุณสมบัติที่ได้รับจากพ่อแม่ในช่วงชีวิต รากเหง้าของมุมมองของดาร์วินเกี่ยวกับกลไกการถ่ายทอดลักษณะจากพ่อแม่สู่ลูกหลานโดยทางสายเลือดอยู่ในปรัชญาธรรมชาติของนักปรัชญากรีกโบราณ รวมถึงคำสอนของฮิปโปเครติส (ศตวรรษที่ 5 ก่อนคริสตกาล)
สมมติฐานเก็งกำไรเรื่องพันธุกรรมอีกข้อหนึ่งถูกหยิบยกขึ้นมาในปี พ.ศ. 2427 โดย K. Nägeli (ชาวเยอรมัน) เขาแนะนำว่าสารพิเศษของการถ่ายทอดทางพันธุกรรมมีส่วนร่วมในการถ่ายทอดความโน้มเอียงทางพันธุกรรมไปยังลูกหลาน - ไอดิโอพลาสซึมซึ่งประกอบด้วยโมเลกุลที่รวบรวมในเซลล์เป็นโครงสร้างคล้ายเกลียวขนาดใหญ่ - ไมเซลล์ ไมเซลล์เชื่อมต่อกันเป็นมัดและสร้างเครือข่ายที่แทรกซึมทุกเซลล์ Idioplasma มีอยู่ในเซลล์สืบพันธุ์และเซลล์ร่างกาย ไซโตพลาสซึมที่เหลือไม่ได้มีส่วนร่วมในการถ่ายทอดคุณสมบัติทางพันธุกรรม อย่างไรก็ตาม สมมติฐานของ K. Nägeli ไม่ได้รับการสนับสนุนจากข้อเท็จจริง คาดการณ์ข้อมูลเกี่ยวกับการมีอยู่และโครงสร้างของสารพาหะทางพันธุกรรม
ก. ไวส์แมนเป็นคนแรกที่ชี้ให้เห็นว่าโครโมโซมเป็นพาหะของพันธุกรรม ในทฤษฎีของเขา เขาสรุปจากข้อสรุปของนักวิทยาวิทยาชาวเยอรมัน วิลเฮล์ม รูซ์ (ค.ศ. 1883) เกี่ยวกับการจัดเรียงเชิงเส้นของปัจจัยทางพันธุกรรม (เมล็ดโครมาติน) ในโครโมโซมและการแยกโครโมโซมตามยาวระหว่างการแบ่งเนื่องจากเป็นวิธีที่เป็นไปได้ในการกระจายเนื้อหาทางพันธุกรรม ทฤษฎี "พลาสซึมของเชื้อโรค" โดย A. Weissman ได้รับรูปแบบสุดท้ายในปี พ.ศ. 2435 เขาเชื่อว่าในสิ่งมีชีวิตมีสารพิเศษของการถ่ายทอดทางพันธุกรรม - "พลาสซึมของเชื้อโรค" วัสดุตั้งต้นของเชื้อพันธุกรรมคือโครงสร้างโครมาตินของนิวเคลียสของเซลล์สืบพันธุ์ พลาสซึมของเชื้อโรคนั้นเป็นอมตะ โดยจะถ่ายทอดผ่านเซลล์สืบพันธุ์ไปยังลูกหลาน ในขณะที่ร่างกายของสิ่งมีชีวิต - ตัวเนื้อ - นั้นเป็นสิ่งมีชีวิตที่ต้องตาย เชื้อพันธุกรรมประกอบด้วยอนุภาคที่ไม่ต่อเนื่อง - biophores ซึ่งแต่ละอนุภาคจะกำหนดคุณสมบัติที่แยกจากกันของเซลล์ Biophores ถูกจัดกลุ่มเป็นปัจจัยกำหนด - อนุภาคที่กำหนดความเชี่ยวชาญเฉพาะของเซลล์ ในทางกลับกันจะรวมกันเป็นโครงสร้างที่มีลำดับสูงกว่า (ides) ซึ่งเกิดโครโมโซมขึ้น (ตามคำศัพท์ของ A. Weisman -)
A. Weisman ปฏิเสธความเป็นไปได้ในการสืบทอดทรัพย์สินที่ได้มา แหล่งที่มาของการเปลี่ยนแปลงทางพันธุกรรมตามคำสอนของพระองค์คือเหตุการณ์ที่เกิดขึ้นระหว่างกระบวนการปฏิสนธิ: การสูญเสียข้อมูลบางส่วน (ลดลง) ในช่วงการเจริญเติบโตของเซลล์สืบพันธุ์และการผสมกันของปัจจัยกำหนดของพ่อและแม่นำไปสู่ การปรากฏตัวของคุณสมบัติใหม่ ทฤษฎีของ A. Weisman มีผลกระทบอย่างมากต่อการพัฒนาทางพันธุศาสตร์ ซึ่งเป็นตัวกำหนดทิศทางต่อไปของการวิจัยทางพันธุกรรม
เมื่อต้นศตวรรษที่ยี่สิบ ข้อกำหนดเบื้องต้นที่แท้จริงถูกสร้างขึ้นเพื่อการพัฒนาวิทยาศาสตร์ทางพันธุกรรม การค้นพบกฎของ G. Mendel อีกครั้งในปี 1900 มีบทบาทชี้ขาด นักวิจัยสมัครเล่นชาวเช็ก พระสงฆ์แห่งอารามบรุนน์ เกรเกอร์ เมนเดล ได้กำหนดกฎพื้นฐานของการถ่ายทอดทางพันธุกรรมย้อนกลับไปในปี พ.ศ. 2408 สิ่งนี้เกิดขึ้นได้เนื่องจากการพัฒนาวิธีการทางพันธุกรรมทางวิทยาศาสตร์วิธีแรกซึ่งเรียกว่า "ไฮบริดวิทยา" มันขึ้นอยู่กับระบบการผสมข้ามพันธุ์ซึ่งทำให้สามารถเปิดเผยรูปแบบการสืบทอดลักษณะได้ เมนเดลได้กำหนดกฎสามข้อและกฎของ "ความบริสุทธิ์ของกาเมต" ซึ่งจะกล่าวถึงรายละเอียดในการบรรยายครั้งต่อไป สิ่งที่สำคัญไม่น้อย (และอาจจะมากกว่านั้น) ก็คือเมนเดลได้แนะนำแนวคิดเกี่ยวกับความโน้มเอียงทางพันธุกรรม (ต้นแบบของยีน) ซึ่งทำหน้าที่เป็นพื้นฐานที่สำคัญสำหรับการพัฒนาลักษณะต่างๆ และแสดงการคาดเดาที่ยอดเยี่ยมเกี่ยวกับการจับคู่ของพวกเขาอันเป็นผลมาจากการผสมผสานของ gametes "บริสุทธิ์"
การวิจัยของเมนเดลและมุมมองของเขาเกี่ยวกับกลไกการถ่ายทอดทางพันธุกรรมนั้นล้ำหน้าการพัฒนาวิทยาศาสตร์ไปหลายทศวรรษ แม้แต่สมมติฐานเชิงเก็งกำไรเกี่ยวกับธรรมชาติของพันธุกรรมที่กล่าวถึงข้างต้นก็ยังได้รับการกำหนดในภายหลัง โครโมโซมยังไม่ถูกค้นพบ และกระบวนการแบ่งเซลล์ซึ่งรองรับการถ่ายทอดข้อมูลทางพันธุกรรมจากพ่อแม่ไปยังลูกหลานนั้น ยังไม่ได้รับการอธิบาย ในเรื่องนี้ผู้ร่วมสมัยแม้กระทั่งผู้ที่คุ้นเคยกับผลงานของ G. Mendel เช่นเดียวกับ Charles Darwin ก็ล้มเหลวในการชื่นชมการค้นพบของเขา เป็นเวลา 35 ปีแล้วที่วิทยาศาสตร์ชีวภาพไม่เป็นที่ต้องการ
ความยุติธรรมได้รับชัยชนะในปี 1900 เมื่อมีการค้นพบกฎของเมนเดลครั้งที่สองอีกครั้งพร้อมกันและเป็นอิสระโดยนักวิทยาศาสตร์สามคน: G. de Vries (ชาวดัตช์), K. Correns (ชาวเยอรมัน) และ E. Cermak (ชาวออสเตรีย) ด้วยการทดลองซ้ำของเมนเดล พวกเขายืนยันลักษณะสากลของรูปแบบที่เขาค้นพบ เมนเดลถือเป็นผู้ก่อตั้งพันธุศาสตร์ และการพัฒนาวิทยาศาสตร์นี้เริ่มขึ้นในปี 1900
ในประวัติศาสตร์ของพันธุศาสตร์ โดยปกติจะแบ่งช่วงเวลาออกเป็นสองช่วง ช่วงแรกคือช่วงของพันธุศาสตร์คลาสสิกหรือเป็นทางการ (ค.ศ. 1900-1944) และช่วงที่สองคือช่วงของอณูพันธุศาสตร์ ซึ่งดำเนินมาจนถึงปัจจุบัน ลักษณะสำคัญของช่วงแรกคือธรรมชาติของสารพาหะทางพันธุกรรมยังไม่ทราบแน่ชัด แนวคิดเรื่อง "ยีน" ซึ่งเป็นอะนาล็อกของปัจจัยทางพันธุกรรม Mendelian ซึ่งแนะนำโดยนักพันธุศาสตร์ชาวเดนมาร์ก V. Johansen นั้นเป็นนามธรรม ต่อไปนี้เป็นคำพูดจากผลงานของเขาในปี 1909: “คุณสมบัติของสิ่งมีชีวิตถูกกำหนดโดยพิเศษภายใต้สถานการณ์บางอย่าง ซึ่งแยกออกจากกัน ดังนั้นในระดับหนึ่งจะมีหน่วยหรือองค์ประกอบอิสระในเซลล์สืบพันธุ์ ซึ่งเราเรียกว่ายีน ในปัจจุบัน เป็นไปไม่ได้ที่จะสร้างแนวคิดที่ชัดเจนเกี่ยวกับธรรมชาติของยีน เราทำได้เพียงพอใจกับข้อเท็จจริงที่ว่าองค์ประกอบดังกล่าวมีอยู่จริงเท่านั้น แต่พวกมันเป็นองค์ประกอบทางเคมีหรือเปล่า? เรายังไม่รู้อะไรเกี่ยวกับเรื่องนี้เลย” แม้จะขาดความรู้เกี่ยวกับธรรมชาติเคมีกายภาพของยีน แต่ในช่วงเวลานี้เองที่มีการค้นพบกฎพื้นฐานของพันธุศาสตร์และทฤษฎีทางพันธุกรรมได้รับการพัฒนาซึ่งเป็นรากฐานของวิทยาศาสตร์นี้
การค้นพบกฎของเมนเดลอีกครั้งในปี ค.ศ. 1900 นำไปสู่การเผยแพร่คำสอนของเขาอย่างรวดเร็ว และความพยายามจำนวนมากที่ประสบความสำเร็จมากที่สุดโดยนักวิจัยในประเทศต่างๆ เกี่ยวกับวัตถุต่างๆ (ไก่ ผีเสื้อ สัตว์ฟันแทะ ฯลฯ) เพื่อยืนยันธรรมชาติสากลของกฎของเขา ในระหว่างการทดลองเหล่านี้ มีการเปิดเผยรูปแบบการสืบทอดแบบใหม่ ในปี 1906 นักวิทยาศาสตร์ชาวอังกฤษ ดับบลิว. แบทสัน และอาร์. ปุนเน็ตต์ บรรยายถึงกรณีแรกของการเบี่ยงเบนจากกฎของเมนเดล ซึ่งต่อมาเรียกว่าการเชื่อมโยงของยีน ในปีเดียวกันนั้น L. Doncaster นักพันธุศาสตร์ชาวอังกฤษในการทดลองกับผีเสื้อได้ค้นพบปรากฏการณ์การเชื่อมโยงระหว่างลักษณะกับเพศ ในเวลาเดียวกันในช่วงต้นศตวรรษที่ยี่สิบ การศึกษาการเปลี่ยนแปลงทางพันธุกรรมอย่างต่อเนื่องในการกลายพันธุ์เริ่มต้นขึ้น (G. de Vries, S. Korzhinsky) และผลงานชิ้นแรกเกี่ยวกับพันธุศาสตร์ประชากรก็ปรากฏขึ้น ในปี 1908 G. Hardy และ V. Weinberg ได้กำหนดกฎพื้นฐานของพันธุศาสตร์ประชากรเกี่ยวกับความคงที่ของความถี่ของยีน
แต่การศึกษาที่สำคัญที่สุดในช่วงเวลาของพันธุศาสตร์คลาสสิกคือผลงานของนักพันธุศาสตร์ชาวอเมริกันผู้มีชื่อเสียง T. Morgan และนักเรียนของเขา ที. มอร์แกนเป็นผู้ก่อตั้งและผู้นำโรงเรียนพันธุศาสตร์ที่ใหญ่ที่สุดในโลกซึ่งมีนักพันธุศาสตร์ที่มีความสามารถทั้งกาแล็กซีเกิดขึ้น ในการวิจัยของเขา มอร์แกนเป็นคนแรกที่ใช้แมลงวันผลไม้ดรอสโซฟิล่า ซึ่งต่อมาได้กลายเป็นวัตถุทางพันธุกรรมที่ชื่นชอบและยังคงเป็นเช่นนี้มาจนถึงทุกวันนี้ การศึกษาปรากฏการณ์การเชื่อมโยงของยีนที่ค้นพบโดย W. Betson และ R. Punnett ทำให้มอร์แกนสามารถกำหนดหลักการพื้นฐานของทฤษฎีพันธุกรรมของโครโมโซมซึ่งเราจะตรวจสอบในรายละเอียดด้านล่าง วิทยานิพนธ์หลักของทฤษฎีทางพันธุกรรมพื้นฐานนี้คือ ยีนถูกจัดเรียงเป็นเส้นตรงบนโครโมโซม เหมือนเม็ดบีดบนเชือก อย่างไรก็ตาม แม้กระทั่งในปี 1937 มอร์แกนเขียนว่าไม่มีข้อตกลงระหว่างนักพันธุศาสตร์เกี่ยวกับธรรมชาติของยีน ไม่ว่าพวกมันจะเป็นของจริงหรือเป็นนามธรรมก็ตาม แต่เขาตั้งข้อสังเกตว่าไม่ว่าในกรณีใด ยีนนั้นมีความเกี่ยวข้องกับโครโมโซมจำเพาะ และสามารถระบุได้เฉพาะที่นั้นผ่านการวิเคราะห์ทางพันธุกรรมล้วนๆ
มอร์แกนและเพื่อนร่วมงานของเขา (T. Paynter, K. Bridges, A. Sturtevant ฯลฯ ) ได้ทำการศึกษาที่โดดเด่นอื่น ๆ อีกมากมาย: หลักการของการทำแผนที่ทางพันธุกรรมได้รับการพัฒนา, ทฤษฎีโครโมโซมของการกำหนดเพศถูกสร้างขึ้นและโครงสร้างของ ศึกษาโครโมโซมโพลีทีน
เหตุการณ์สำคัญในช่วงเวลาของพันธุศาสตร์คลาสสิกคือการพัฒนางานเกี่ยวกับการกลายพันธุ์เทียมซึ่งเป็นข้อมูลแรกที่ได้รับในปี 1925 ในสหภาพโซเวียตโดย G.A. แนดสันและที.เอส. Filippov ในการทดลองการฉายรังสีเซลล์ยีสต์ด้วยเรเดียม สิ่งสำคัญอย่างยิ่งสำหรับการพัฒนางานในทิศทางนี้คือการทดลองของนักพันธุศาสตร์ชาวอเมริกัน G. Meller เกี่ยวกับผลกระทบของรังสีเอกซ์ต่อดรอสโซฟิล่าและการพัฒนาวิธีการบันทึกการกลายพันธุ์ในเชิงปริมาณ งานของ G. Möller ก่อให้เกิดการศึกษาทดลองจำนวนมากโดยใช้รังสีเอกซ์บนวัตถุต่างๆ เป็นผลให้เกิดผลกระทบต่อการกลายพันธุ์ที่เป็นสากล ต่อมาได้ค้นพบว่ารังสีชนิดอื่น เช่น รังสียูวี รวมถึงอุณหภูมิสูงและสารเคมีบางชนิดก็มีผลกระทบต่อการกลายพันธุ์เช่นกัน สารก่อกลายพันธุ์ทางเคมีชนิดแรกถูกค้นพบในช่วงทศวรรษที่ 30 ในสหภาพโซเวียตในการทดลองของ V.V. Sakharova, M.E. Lobashev และ S.M. เกอร์เชนซอนและพนักงานของพวกเขา ไม่กี่ปีต่อมา ทิศทางนี้ได้รับขอบเขตกว้างขวาง โดยเฉพาะอย่างยิ่งต้องขอบคุณการวิจัยของ A.I. Rapoport ในสหภาพโซเวียตและ S. Auerbach ในอังกฤษ
การวิจัยในสาขาการทดลองก่อกลายพันธุ์ได้นำไปสู่ความก้าวหน้าอย่างรวดเร็วในการทำความเข้าใจกระบวนการกลายพันธุ์ และการชี้แจงประเด็นต่างๆ ที่เกี่ยวข้องกับโครงสร้างที่ดีของยีน
การวิจัยทางพันธุกรรมที่สำคัญอีกประการหนึ่งในช่วงพันธุศาสตร์คลาสสิกเกี่ยวข้องกับการศึกษาบทบาทของกระบวนการทางพันธุกรรมในวิวัฒนาการ ผลงานสำคัญในพื้นที่นี้เป็นของ S. Wright, R. Fisher, J. Haldane และ S.S. เชตเวริคอฟ. ด้วยผลงานของพวกเขา พวกเขายืนยันความถูกต้องของหลักการพื้นฐานของลัทธิดาร์วินและมีส่วนทำให้เกิดทฤษฎีวิวัฒนาการสังเคราะห์สมัยใหม่ใหม่ ซึ่งเป็นผลมาจากการสังเคราะห์ทฤษฎีของดาร์วินและพันธุศาสตร์ประชากร
ตั้งแต่ปีพ. ศ. 2483 ช่วงที่สองของการพัฒนาพันธุศาสตร์โลกเริ่มขึ้นซึ่งเรียกว่าโมเลกุลตามตำแหน่งผู้นำของทิศทางของพันธุศาสตร์นี้ บทบาทหลักในการเพิ่มขึ้นอย่างรวดเร็วของอณูพันธุศาสตร์แสดงโดยพันธมิตรอย่างใกล้ชิดของนักชีววิทยากับนักวิทยาศาสตร์จากสาขาวิทยาศาสตร์ธรรมชาติอื่นๆ (ฟิสิกส์ คณิตศาสตร์ ไซเบอร์เนติกส์ เคมี) ภายหลังจากที่มีการค้นพบที่สำคัญหลายประการ ในช่วงเวลานี้ นักวิทยาศาสตร์ได้กำหนดลักษณะทางเคมีของยีน กำหนดกลไกการออกฤทธิ์และการควบคุม และค้นพบที่สำคัญอีกมากมายที่เปลี่ยนพันธุกรรมให้เป็นหนึ่งในสาขาวิชาทางชีววิทยาหลักที่กำหนดความก้าวหน้าของวิทยาศาสตร์ธรรมชาติสมัยใหม่ การค้นพบอณูพันธุศาสตร์ไม่ได้ปฏิเสธ แต่เพียงเผยให้เห็นกลไกเชิงลึกของรูปแบบทางพันธุกรรมเหล่านั้นที่นักพันธุศาสตร์อย่างเป็นทางการเปิดเผยเท่านั้น
ผลงานของ J. Beadle และ E. Tetum (สหรัฐอเมริกา) พบว่าการกลายพันธุ์ในเชื้อรา Neurospora crassa ของเชื้อราขนมปังขัดขวางขั้นตอนต่างๆ ของการเผาผลาญของเซลล์ ผู้เขียนแนะนำว่ายีนควบคุมการสังเคราะห์ทางชีวภาพของเอนไซม์ วิทยานิพนธ์เกิดขึ้น: “หนึ่งยีน หนึ่งเอนไซม์” ในปี 1944 การศึกษาเกี่ยวกับการเปลี่ยนแปลงทางพันธุกรรมในแบคทีเรียดำเนินการโดยนักวิทยาศาสตร์ชาวอเมริกัน (O. Avery, K. McLeod และ M. McCarthy) แสดงให้เห็นว่า DNA เป็นพาหะของข้อมูลทางพันธุกรรม ข้อสรุปนี้ได้รับการยืนยันในภายหลังโดยการศึกษาปรากฏการณ์การถ่ายโอน (J. Lederberg และ M. Zinder, 1952) ซึ่งเป็นการถ่ายโอนข้อมูลจากเซลล์แบคทีเรียหนึ่งไปยังอีกเซลล์หนึ่งโดยใช้ phage DNA
การศึกษาที่ระบุไว้ได้กำหนดความสนใจที่เพิ่มขึ้นในการศึกษาโครงสร้างของ DNA ซึ่งส่งผลให้เกิดการสร้างแบบจำลองโมเลกุล DNA ในปี 1953 โดย J. Watson (นักชีววิทยาชาวอเมริกัน) และ F. Crick (นักเคมีชาวอังกฤษ) มันถูกเรียกว่าเกลียวคู่เพราะตามแบบจำลอง มันถูกสร้างขึ้นจากสายพอลินิวคลีโอไทด์สองเส้นที่บิดเป็นเกลียว DNA เป็นโพลีเมอร์ที่มีโมโนเมอร์เป็นนิวคลีโอไทด์ นิวคลีโอไทด์แต่ละตัวประกอบด้วยดีออกซีไรโบสน้ำตาลห้าคาร์บอน กรดฟอสฟอริกตกค้าง และเบสไนโตรเจนหนึ่งในสี่เบส (อะดีนีน กัวนีน ไซโตซีน และไทมีน) งานนี้มีบทบาทสำคัญในการพัฒนาต่อไปด้านพันธุศาสตร์และอณูชีววิทยา
จากแบบจำลองนี้ กลไกกึ่งอนุรักษ์นิยมของการสังเคราะห์ DNA ได้รับการตั้งสมมติฐานครั้งแรก (F. Crick) จากนั้นจึงพิสูจน์เชิงทดลอง (M. Meselson และ F. Stahl, 1957) ซึ่งโมเลกุล DNA แบ่งออกเป็นสองสายเดี่ยวแต่ละสาย ซึ่งทำหน้าที่เป็นเทมเพลตสำหรับการสังเคราะห์ห่วงโซ่ลูก การสังเคราะห์ขึ้นอยู่กับหลักการของการเสริมซึ่งกันและกัน ซึ่งกำหนดไว้ก่อนหน้านี้โดย E. Chargaff (1945) โดยที่ฐานไนโตรเจนของสาย DNA สองสายตั้งอยู่ตรงข้ามกันเป็นคู่ โดยมีอะดีนีนเชื่อมต่อเฉพาะกับไทมีน (A-T) และกัวนีน ไปสู่ไซโตซีน (G-C) ผลที่ตามมาประการหนึ่งของการสร้างแบบจำลองคือการถอดรหัสรหัสพันธุกรรม - หลักการบันทึกข้อมูลทางพันธุกรรม ทีมวิทยาศาสตร์จำนวนมากในประเทศต่างๆ ได้แก้ไขปัญหานี้แล้ว ความสำเร็จมาถึงอาเมอร์ นักพันธุศาสตร์ M. Nirenberg (ผู้ได้รับรางวัลโนเบล) ซึ่งถอดรหัสคำรหัสแรกในห้องปฏิบัติการคือ codon คำนี้กลายเป็นแฝดสาม YYY ซึ่งเป็นลำดับของนิวคลีโอไทด์สามตัวที่มีฐานไนโตรเจนเหมือนกัน - ยูราซิล เมื่อมีโมเลกุล mRNA ซึ่งประกอบด้วยสายโซ่ของนิวคลีโอไทด์ดังกล่าว โปรตีนที่จำเจจะถูกสังเคราะห์ซึ่งมีสารตกค้างที่เชื่อมต่อกันตามลำดับของกรดอะมิโนชนิดเดียวกัน - ฟีนิลอะลานีน การถอดรหัสรหัสเพิ่มเติมเป็นเรื่องของเทคโนโลยี: นักวิทยาศาสตร์ได้รวบรวมตารางรหัสโดยใช้เมทริกซ์ที่มีฐานที่แตกต่างกันในรหัส คุณสมบัติทั้งหมดของรหัสพันธุกรรมได้รับการพิจารณา: ความเป็นสากล, ความเป็นสามเท่า, ความเสื่อมและการไม่ทับซ้อนกัน การถอดรหัสรหัสพันธุกรรมในแง่ของความสำคัญต่อการพัฒนาวิทยาศาสตร์และการปฏิบัตินั้นเปรียบเทียบกับการค้นพบพลังงานนิวเคลียร์ในฟิสิกส์
หลังจากถอดรหัสรหัสพันธุกรรมและกำหนดหลักการบันทึกข้อมูลทางพันธุกรรมแล้ว นักวิทยาศาสตร์ก็เริ่มคิดว่าข้อมูลถูกถ่ายโอนจาก DNA ไปยังโปรตีนได้อย่างไร การวิจัยปัญหานี้จบลงด้วยคำอธิบายที่สมบูรณ์เกี่ยวกับกลไกในการรับทราบข้อมูลทางพันธุกรรม ซึ่งรวมถึงสองขั้นตอน: การถอดความและการแปล
หลังจากพิจารณาลักษณะทางเคมีของยีนและหลักการทำงานของยีนแล้ว คำถามก็เกิดขึ้นว่าควบคุมการทำงานของยีนอย่างไร ได้ยินครั้งแรกในการศึกษาของนักชีวเคมีชาวฝรั่งเศส F. Jacob และ J. Monod (1960) ซึ่งเป็นผู้พัฒนาโครงการควบคุมกลุ่มยีนที่ควบคุมกระบวนการหมักแลคโตสในเซลล์ E. coli พวกเขานำเสนอแนวคิดของโอเปอเรเตอร์ของแบคทีเรียในฐานะคอมเพล็กซ์ที่รวมยีนทั้งหมด (ทั้งยีนที่มีโครงสร้างและยีนควบคุม) ที่ให้บริการส่วนใดส่วนหนึ่งของการเผาผลาญ ต่อมาความถูกต้องของโครงการได้รับการพิสูจน์โดยการทดลองโดยการศึกษาการกลายพันธุ์ต่างๆ ที่ส่งผลต่อหน่วยโครงสร้างต่างๆ ของโอเปอเรเตอร์
ได้มีการพัฒนาแผนสำหรับกลไกการควบคุมยีนยูคาริโอตอย่างค่อยเป็นค่อยไป สิ่งนี้ได้รับการอำนวยความสะดวกโดยการสร้างโครงสร้างที่ไม่ต่อเนื่องของยีนบางตัวและคำอธิบายของกลไกการต่อรอย
ได้รับอิทธิพลจากความก้าวหน้าในการศึกษาโครงสร้างและหน้าที่ของยีนในช่วงต้นทศวรรษที่ 70 ศตวรรษที่ XX นักพันธุศาสตร์เกิดแนวคิดในการจัดการกับพวกมันก่อนอื่นโดยการถ่ายโอนพวกมันจากเซลล์หนึ่งไปอีกเซลล์หนึ่ง นี่คือลักษณะที่ทิศทางใหม่ของการวิจัยทางพันธุกรรมปรากฏขึ้น - พันธุวิศวกรรม
พื้นฐานสำหรับการพัฒนาทิศทางนี้เกิดขึ้นจากการทดลองในระหว่างที่มีการพัฒนาวิธีการรับยีนแต่ละตัว ในปี 1969 ในห้องปฏิบัติการของ J. Beckwith โอเปอเรเตอร์แลคโตสถูกแยกออกจากโครโมโซม E. coli โดยใช้ปรากฏการณ์การถ่ายโอน ในปี 1970 ทีมงานที่นำโดย G. Corano ทำการสังเคราะห์ทางเคมีของยีนเป็นครั้งแรก ในปี 1973 ได้มีการพัฒนาวิธีการผลิตชิ้นส่วน DNA ซึ่งเป็นผู้บริจาคยีน โดยใช้เอนไซม์จำกัดที่จะตัดโมเลกุล DNA และในที่สุดวิธีการรับยีนได้รับการพัฒนาตามปรากฏการณ์ของการถอดรหัสแบบย้อนกลับซึ่งค้นพบในปี 1975 โดย D. Baltimore และ G. Temin เพื่อนำยีนแปลกปลอมเข้าสู่เซลล์ พาหะต่างๆ—โมเลกุลพาหะ—ที่ดำเนินการกระบวนการถ่ายโอนถูกสร้างขึ้นบนพื้นฐานของพลาสมิด ไวรัส แบคทีริโอฟาจ และทรานสโพซัน (องค์ประกอบทางพันธุกรรมที่เคลื่อนที่ได้) คอมเพล็กซ์เวกเตอร์-ยีนถูกเรียกว่าโมเลกุลรีคอมบิแนนท์ โมเลกุลที่มีพื้นฐานจากดีเอ็นเอฟาจรีคอมบิแนนท์ตัวแรกถูกสร้างขึ้นในปี 1974 (อาร์. เมอร์เรย์และดี. เมอร์เรย์) ในปี พ.ศ. 2518 ได้มีการพัฒนาวิธีการสำหรับการโคลนเซลล์และฟาจที่มียีนแทรกอยู่
แล้วในช่วงต้นทศวรรษที่ 70 ได้รับผลการทดลองครั้งแรกในสาขาพันธุวิศวกรรม ดังนั้นโมเลกุลรีคอมบิแนนท์ที่มียีนต้านทานยาปฏิชีวนะสองตัว (เตตราไซคลินและสเตรปโตมัยซิน) จึงถูกนำเข้าไปในเซลล์ E. coli หลังจากนั้นเซลล์ก็ต้านทานยาทั้งสองชนิด
ชุดของเวกเตอร์และยีนที่แนะนำค่อยๆ ขยายตัวและเทคโนโลยีการถ่ายโอนได้รับการปรับปรุง สิ่งนี้ทำให้เป็นไปได้ที่จะใช้วิธีการทางพันธุวิศวกรรมเพื่อวัตถุประสงค์ทางอุตสาหกรรม (เทคโนโลยีชีวภาพ) อย่างกว้างขวาง โดยเน้นไปที่ผลประโยชน์ด้านการแพทย์และการเกษตรเป็นหลัก แบคทีเรียได้รับการออกแบบเพื่อผลิตสารออกฤทธิ์ทางชีวภาพ สิ่งนี้ทำให้สามารถสร้างการสังเคราะห์ยาที่จำเป็นสำหรับมนุษย์ในระดับที่ต้องการเช่นอินซูลิน, โซมาโตสตาติน, อินเตอร์เฟอรอน, ทริปโตเฟน ฯลฯ มีการสร้างพืชดัดแปรพันธุกรรมจำนวนมากซึ่งกลายเป็นเจ้าของคุณสมบัติที่มีคุณค่า (ความต้านทานต่อศัตรูพืชความแห้งแล้ง ปริมาณโปรตีนสูง ฯลฯ) ซึ่งเป็นผลมาจากการนำยีนจากต่างประเทศเข้าสู่จีโนม
ในยุค 70 งานเริ่มต้นจากการจัดลำดับจีโนมของวัตถุต่างๆ ตั้งแต่แบคทีเรียไปจนถึงมนุษย์
โปรแกรมพันธุกรรมระหว่างประเทศ "จีโนมมนุษย์" สมควรได้รับความสนใจเป็นพิเศษ โดยมีเป้าหมายเพื่อถอดรหัสรหัสพันธุกรรมของมนุษย์อย่างสมบูรณ์และทำแผนที่โครโมโซมของมัน ในอนาคตมีการวางแผนการพัฒนาอย่างเข้มข้นของพันธุศาสตร์การแพทย์สาขาใหม่ - การบำบัดด้วยยีนซึ่งจะช่วยลดความเสี่ยงของการแสดงออกของยีนที่เป็นอันตรายและด้วยเหตุนี้จึงจำกัดภาระทางพันธุกรรมให้สูงสุด
การก่อตัวของพันธุศาสตร์ในรัสเซียเกิดขึ้นในทศวรรษที่สองของศตวรรษที่ยี่สิบ ผู้ก่อตั้งโรงเรียนนักพันธุศาสตร์ในประเทศแห่งแรกคือ Yuri Aleksandrovich Filipchenko ในปี 1916 เขาเริ่มบรรยายหลักสูตร "The Doctrine of Heredity and Evolution" ที่มหาวิทยาลัยเซนต์ปีเตอร์สเบิร์ก ซึ่งเขาได้เป็นศูนย์กลางของกฎของ Mendel และการวิจัยของ T. Morgan เขาแปลหนังสือ "The Gene Theory" ของมอร์แกนโดยได้รับอนุญาต ความสนใจทางวิทยาศาสตร์ของ Yu.A. Filipchenko อยู่ในแวดวงพันธุกรรมและความแปรปรวนของลักษณะเชิงคุณภาพและเชิงปริมาณ เขาให้ความสนใจเป็นพิเศษกับรูปแบบทางสถิติของความแปรปรวน ยุเอ Filipchenko เขียนหนังสือที่ยอดเยี่ยมหลายเล่ม หนึ่งในนั้นคือตำราเรียนเรื่อง "พันธุศาสตร์" ซึ่งนักชีววิทยาหลายรุ่นศึกษาในประเทศของเรา
ในช่วงเวลาเดียวกันมีการจัดตั้งโรงเรียนพันธุศาสตร์วิทยาศาสตร์อีกสองแห่ง: แห่งหนึ่งที่สถาบันชีววิทยาทดลอง (มอสโก) ภายใต้การนำของ Nikolai Konstantinovich Koltsov และอีกแห่งภายใต้การนำของ Nikolai Ivanovich Vavilov เริ่มสร้างขึ้นใน Saratov ซึ่งเขาอยู่ ได้รับเลือกเป็นอาจารย์มหาวิทยาลัย และในที่สุดก็ก่อตั้งขึ้นในเลนินกราดบนพื้นฐานของ All-Union Institute of Plant Growing (VIR)
เอ็น.เค. Koltsov เป็นหัวหน้าสถาบันวิจัยชีววิทยาทดลองขนาดใหญ่ในมอสโก เขาเป็นคนแรกที่แสดงความคิดเกี่ยวกับการจัดระเบียบโมเลกุลขนาดใหญ่ของพาหะของพันธุกรรม (โครโมโซม) และการทำซ้ำตัวเองเป็นกลไกในการส่งข้อมูลทางพันธุกรรม ไอเดีย เอ็น.เค. Koltsov มีอิทธิพลอย่างมากต่อนักวิทยาศาสตร์ที่มีชื่อเสียงในยุคนั้น ไม่เพียงแต่นักชีววิทยาเท่านั้น แต่ยังรวมถึงนักฟิสิกส์ด้วย ซึ่งการศึกษาโครงสร้างยีนนำไปสู่การพัฒนาพันธุศาสตร์ระดับโมเลกุล จากโรงเรียนวิทยาศาสตร์ของ N.K. Koltsov รวมนักพันธุศาสตร์ที่มีชื่อเสียงเช่น A.S. เซเรบรอฟสกี้ บี.แอล. อัสเตารอฟ, N.P. ดูบินิน, N.V. Timofeev-Resovsky, V.V. ซาคารอฟและคนอื่นๆ
นักพันธุศาสตร์และผู้เพาะพันธุ์ดีเด่น N.I. Vavilov ได้รับการยอมรับอย่างกว้างขวางจากผลงานของเขาในการศึกษาการเกษตรและทรัพยากรพืชโลก เขาเป็นผู้เขียนหลักคำสอนเกี่ยวกับศูนย์กลางของแหล่งกำเนิดและความหลากหลายของพืชที่ปลูก และหลักคำสอนเรื่องภูมิคุ้มกัน เช่นเดียวกับกฎของอนุกรมที่คล้ายคลึงกันในความแปรปรวนทางพันธุกรรม นอกจากนี้ เขายังได้สร้างคอลเลคชันพืชทางการเกษตรและเทคนิคระดับโลก รวมถึงคอลเลคชันข้าวสาลีพันธุ์ที่มีชื่อเสียงอีกด้วย เอ็นไอ วาวิลอฟมีความสุขกับอำนาจอันยิ่งใหญ่ไม่เพียงแต่ในหมู่นักวิทยาศาสตร์ในประเทศเท่านั้น แต่ยังรวมถึงนักวิทยาศาสตร์ชาวต่างชาติด้วย นักวิทยาศาสตร์จากทั่วทุกมุมโลกมาทำงานที่ All-Union Institute of Plant Growing (VIR) ที่เขาสร้างขึ้นในเลนินกราด การรับรู้ถึงข้อดีของ N.I. Vavilov ได้รับเลือกเป็นประธานของ International Genetic Congress ซึ่งจัดขึ้นในปี 1937 ที่เมืองเอดินบะระ อย่างไรก็ตาม สถานการณ์ไม่อนุญาตให้ N.I. Vavilov เข้าร่วมการประชุมครั้งนี้
การวิจัยของศาสตราจารย์ Alexander Sergeevich Serebrovsky จากมหาวิทยาลัยมอสโกและเพื่อนร่วมงานรุ่นเยาว์ของเขา N.P. ดูบินีนา บี.เอ็น. ซิโดโรวา, I.I. อาโกล และคนอื่นๆ. ในปีพ. ศ. 2472 พวกเขาได้ค้นพบปรากฏการณ์ของอัลลีลิสแบบขั้นตอนในดรอสโซฟิล่าซึ่งกลายเป็นก้าวแรกในการละทิ้งแนวคิดเรื่องการแบ่งแยกยีนซึ่งได้กลายมาเป็นที่ยอมรับในหมู่นักพันธุศาสตร์. ทฤษฎีศูนย์กลางของโครงสร้างยีนได้รับการกำหนดขึ้น โดยยีนประกอบด้วยหน่วยย่อยที่เล็กกว่า ซึ่งเป็นศูนย์กลางที่สามารถกลายพันธุ์โดยอิสระจากกัน การศึกษาเหล่านี้เป็นแรงผลักดันในการพัฒนางานเกี่ยวกับการศึกษาโครงสร้างและหน้าที่ของยีนซึ่งส่งผลให้เกิดการพัฒนาแนวคิดสมัยใหม่ของการจัดระเบียบภายในที่ซับซ้อนของยีน ต่อมา (ในปี พ.ศ. 2509) สำหรับผลงานชุดหนึ่งในสาขาทฤษฎีการกลายพันธุ์ N.P. Dubinin ได้รับรางวัลเลนิน
เมื่อต้นทศวรรษที่ 40 ศตวรรษที่ XX ในสหภาพโซเวียต พันธุศาสตร์อยู่ในช่วงรุ่งเรือง นอกเหนือจากที่กล่าวมาข้างต้นแล้ว ควรสังเกตผลงานของ B.L. Astaurov เรื่องการควบคุมเพศในหนอนไหมโดยวิธีทางพันธุกรรม การศึกษาทางเซลล์พันธุศาสตร์ G.A. Levitsky ผลงานของ A.A. ซาเปจิน่า เค.เค. ไมสเตอร์, เอ.อาร์. Zhebraka, N.V. Tsitsin เกี่ยวกับพันธุศาสตร์และการปรับปรุงพันธุ์พืช ม.ฟ. Ivanov เกี่ยวกับพันธุศาสตร์และการปรับปรุงพันธุ์สัตว์ วี.วี. Sakharova, M.E. Lobasheva, S.M. เกอร์เชนโซนา ไอ.เอ. รายงานเกี่ยวกับการกลายพันธุ์ทางเคมี เอส.จี. เลวิต และ S.N. Davidenkov เกี่ยวกับพันธุศาสตร์มนุษย์และผลงานของนักวิทยาศาสตร์ผู้มีความสามารถคนอื่นๆ อีกมากมาย
อย่างไรก็ตาม สถานการณ์ทางการเมืองของการเผชิญหน้ากับโลกทุนนิยมที่พัฒนาขึ้นในสหภาพโซเวียตในช่วงเริ่มต้นของสงครามโลกครั้งที่สอง นำไปสู่การประหัตประหารนักวิทยาศาสตร์ที่ทำงานในสาขาพันธุศาสตร์ ซึ่งได้รับการประกาศให้เป็นวิทยาศาสตร์ชนชั้นกลางในอุดมคติ และสมัครพรรคพวกถูกประกาศให้เป็นตัวแทนของ ของจักรวรรดินิยมโลก การปราบปรามตกอยู่บนหัวของนักวิทยาศาสตร์ชื่อดังหลายคนรวมถึง N.I. วาวิโลวา มิ.ย. Lobasheva, G.D. คาร์เปเชนโก, S.M. Gershenzon และคนอื่นๆ อีกมากมาย พันธุศาสตร์ย้อนกลับไปหลายทศวรรษแล้ว T.D. มีบทบาทสำคัญในการล่มสลายของพันธุศาสตร์ ลีเซนโก. เนื่องจากเป็นนักปฐพีวิทยาธรรมดาๆ เขาจึงไม่สามารถก้าวไปสู่ระดับพันธุกรรมคลาสสิกด้วยแนวคิดเชิงนามธรรมเกี่ยวกับยีนได้ ดังนั้นเขาจึงปฏิเสธกฎของเมนเดล ทฤษฎีโครโมโซมทางพันธุกรรมของมอร์แกน และหลักคำสอนเรื่องการกลายพันธุ์ Lysenko ปกปิดความไม่สอดคล้องทางวิทยาศาสตร์ของเขาด้วยคำมั่นสัญญาที่ว่าจะเพิ่มขึ้นอย่างรวดเร็วในด้านการเกษตรโดยใช้วิธีการที่เขาสนับสนุนในการเปลี่ยนแปลงพืชภายใต้อิทธิพลของสภาพการเจริญเติบโตซึ่งได้รับการสนับสนุนจาก I.V. สตาลิน Lysenko ใช้ผลงานของผู้เพาะพันธุ์ที่โดดเด่น I.V. มิชูรินา. ไม่เหมือนกับวิทยาศาสตร์โลก พันธุกรรมของเราเริ่มถูกเรียกว่ามิชูริน "เกียรติ" ดังกล่าวนำไปสู่ชื่อเสียงของ Michurin ในฐานะผู้ยึดมั่นในแนวคิดของ Lysenko ซึ่งไม่ได้ละทิ้งนักวิทยาศาสตร์แม้หลังจากการล่มสลายของกิจกรรมของฝ่ายหลังก็ตาม อันที่จริง I.V. มิชูรินเป็นนักเพาะพันธุ์และผู้ปลูกผลไม้ที่ใช้งานได้จริงและไม่เคยมีส่วนเกี่ยวข้องกับการพัฒนารากฐานทางทฤษฎีของพันธุศาสตร์เลย
ในที่สุดวิทยาศาสตร์ในประเทศก็ถูกกำจัดจาก "ลัทธิลิเซนโค" ในช่วงกลางทศวรรษที่ 60 เท่านั้น นักวิทยาศาสตร์หลายคนที่ได้รับความทุกข์ทรมานจากการกดขี่ออกมาจาก "ใต้ดิน" ซึ่งเป็นกลุ่มที่สามารถเอาชีวิตรอดได้รวมถึง N.V. Timofeev-Resovsky, M.E. Lobashov, V.V. ซาคารอฟและคนอื่นๆ ประเพณีที่พวกเขารักษาไว้และศักยภาพอันยิ่งใหญ่ที่มีอยู่ในตัวนักเรียนมีส่วนทำให้เกิดความก้าวหน้าอย่างรวดเร็ว แม้ว่าความล่าช้าในระดับโลกจะทำให้ตัวเองรู้สึกได้ก็ตาม อย่างไรก็ตาม นักพันธุศาสตร์ในประเทศรุ่นใหม่มีจำนวนเพิ่มขึ้น ซึ่งจะต้องนำวิทยาศาสตร์นี้ไปสู่ระดับก่อนหน้า และอีกครั้งที่ชื่อรัสเซียเต็มไปด้วยชื่อนักวิทยาศาสตร์ชื่อดังระดับโลก: A.N. เบโลเซอร์สกี้, เวอร์จิเนีย เองเกลฮาร์ด, S.I. อลิคานยาน, อาร์.บี. Khesina, A.S. สไปรินา เอส.วี. Shestakova, S.G. Inge-Vechtomova, Y.P. Altukhov และคนอื่น ๆ อีกมากมาย
อย่างไรก็ตาม การเปลี่ยนแปลงทางสังคมครั้งใหม่ที่เกิดจากเปเรสทรอยกา ซึ่งนำไปสู่การหลั่งไหลของบุคลากรทางวิทยาศาสตร์ในต่างประเทศ ทำให้วิทยาศาสตร์ของเราไม่ได้รับสถานะที่เหมาะสมอีกครั้ง เราหวังเพียงว่าคนรุ่นใหม่ซึ่งอาศัยรากฐานที่ผู้ทรงคุณวุฒิคนก่อนวางไว้จะสามารถบรรลุภารกิจอันสูงส่งนี้ได้
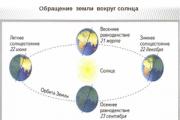
ตั้งแต่สมัยโบราณ ผู้คนสนใจว่าเหตุใดกลางคืนจึงหลีกทางให้กลางวัน ฤดูหนาวในฤดูใบไม้ผลิ และฤดูร้อนในฤดูใบไม้ร่วง ต่อมาเมื่อครั้งแรก...

“ ประวัติศาสตร์ประเทศบ้านเกิดของเรา” อุทิศให้กับวันครบรอบ 100 ปีของการปฏิวัติในรัสเซียในปี พ.ศ. 2460 คำนำถึงต้นศตวรรษที่ 20 ในประเทศที่มีการพัฒนามากที่สุด...

กระต่ายทำเสียงอะไร เกา เสียงคำราม =)) และเมื่อมันโตขึ้น มันก็คำราม! สิ่งเหล่านี้มีหลายแง่มุม...

ชายชราและหญิงชราเป็นคนช่างพูด เทพนิยาย!! .... กาลครั้งหนึ่งมีชายชราและหญิงชราคนหนึ่ง หญิงชราไม่สามารถกลั้นลิ้นของเธอได้...

ย้อนกลับ โปรดทราบ! การแสดงตัวอย่างสไลด์มีวัตถุประสงค์เพื่อให้ข้อมูลเท่านั้นและอาจ...

งาน 19. เครื่องหมายวรรคตอนในประโยคที่ซับซ้อนพร้อมการเชื่อมต่อประเภทต่างๆ ถ้อยคำของงาน: จัดป้าย...

สถานการณ์ เป็นส่วนย่อยของประโยคที่แสดงถึงสัญญาณของการกระทำหรือสัญญาณอื่น....

ชีวประวัติและผลงานของเขาเผยให้เห็นถึงบุคลิกของกบฏ บุคคลที่ไม่คิดว่าตัวเองอยู่ในยุคแห่งความไร้กฎหมายทั่วไป...

ในวันนี้ - 19 กรกฎาคม พ.ศ. 2368 - วันที่ Anna Petrovna ออกจาก Trigorskoye พุชกินให้เธอ...

Nikolai Aleksandrovich Morozov เกิดเมื่อวันที่ 25 มิถุนายน พ.ศ. 2397 บนที่ดิน Borok ในจังหวัด Yaroslavl แม่ของเขาคือ...

คำอธิบายการนำเสนอในแต่ละสไลด์: 1 สไลด์ คำอธิบายของสไลด์: 2 สไลด์ คำอธิบายของสไลด์:...

ในโลกทัศน์ของมนุษยชาติ ในขั้นตอนของการพัฒนานี้ สังคมมีความหลากหลาย มันถูกบังคับ...

ประวัติความเป็นมาของการวิเคราะห์สเปกตรัมอะตอมเริ่มต้นด้วยการทดลองของไอแซก นิวตัน (1666) เกี่ยวกับการสลายตัวของแสงให้เป็นสเปกตรัม....

การศึกษาเพิ่มเติมภาษาอังกฤษระดับประถมศึกษา “การพูดภาษาอังกฤษ” ครู MBOU...

“ประวัติศาสตร์ประเทศบ้านเกิดของเรา” อุทิศให้กับวันครบรอบ 100 ปีของการปฏิวัติในรัสเซีย พ.ศ. 2460 คำนำถึงต้นศตวรรษที่ 20 ใน...

กระต่ายทำเสียงอะไร เกา เสียงคำราม =)) และเมื่อมันโตขึ้น มันก็คำราม! เหล่านี้คือ...